История открытия. Отдельные свойства возбудителя бешенства впервые изучены в 1880 г. Л. Пастером. Внутриклеточные кристаллы вирусов бешенства обнаружены в 1892 г. В. Бабешем и в 1903 г. А. Негри. Первая инактивированная антирабическая вакцина была получена Л. Пастером и сотрудниками в 1885 г. методом аттенуации дикого штамма вируса в мозге восприимчивых животных.
Особенности вируса. Вирион вируса бешенства пулевидной формы, длиной 80-180 нм и шириной 60-80 нм (рис. 2). Суперкапсид липидный, с шипами длиной 7 нм, является производным клетки-хозяина. G-гликопротеины суперкапсида (гемагглютинины) являются протективными антигенами. Геном представлен однониточной линейной несегментированной «-»-РНК. РНК кодирует 5 белков: неструктурный – NS, матриксный – М, гликопротеид – G, РНК-зависимую РНК-полимеразу — L и белок – N, который образует белковые субъединицы нуклеокапсида. Состав вирионов: белки – 72 %, липиды – 22 %, углеводы – 3 %, РНК – 1 %.
Культивирование. Для культивирования вирусов бешенства используют: 1) лабораторных животных (мыши, крысы, обезьяны, собаки, кролики, хомяки); 2) куриные эмбрионы; 3) культуры клеток (опухолевые, Vero, человеческие диплоидные, куриные и утиные эмбриональные клетки).
Резистентность. Вирусы бешенства инактивируются растворами формалина, лизола, фенола, хлорамина, калия перманганата, карболовой кислоты, а также эфиром, ацетоном, спиртом, трипсином, ультрафиолетовыми лучами. При температуре 100˚С вирусы погибают в течение 2 минут, при 60˚С – в течение 10-15 минут.
Эпидемиология. Бешенство – типичный зооноз; от человека к человеку бешенство не передаётся. Резервуаром и источником инфекции являются: 1) псовые (собаки, лисы, волки, енотовидные собаки, шакалы); 2) куньи (куницы, барсуки, ласки, хорьки, горностаи, скунсы, мангусты); 3) рукокрылые (насекомоядные и плотоядные летучие мыши); 4) копытные (коровы, лошади); 5) кошачьи (домашние кошки, дикие кошки, рыси и др.); 6) грызуны (мыши, крысы). Бешенством болеет человек. Механизм передачи инфекции от больного животного к человеку – перкутанный (при укусах животными, либо при ослюнении). Бешенство у собак увеличивается летом. Инкубационный период при бешенстве у животных – 10 дней. Бессимптомного вирусоносительства у животных не бывает.
Патогенез. У животных вирусы бешенства инфицируют головной мозг и слюнные железы. Со слюной вирусы попадают в окружающую среду. В 1 мл слюны бешенного животного содержится 10 8 инфицирующих доз. После укуса или ослюнения раны вирусы бешенства остаются в очаге в течение инкубационного периода, длительность которого варьирует от 12 дней до более года. При связывании с тканью вирус бешенства концентрируется в нервно-мышечных контактах. Тканевым рецептором для вируса бешенства являются ацетилхолиновые и другие рецепторы. Вирусы проникают в аксоны нейронов и попадают в центральную нервную систему. В цитоплазме нейронов гиппокампа, продолговатого мозга, ядер черепно-мозговых нервов, симпатических ганглиев и клетках Пуркинье мозжечка происходит размножение и накопление вирусов бешенства. В пораженных клетках обнаруживаются вирусоспецифические цитоплазматические включения – тельца Бабеша-Негри. Вокруг инфицированных нейронов наблюдаются явления нейронофагии и сателлитоза. Репродукция вирусов бешенства сопровождается увеличением протеина bax и апоптозом нейронов. Поражение нейронов ведет к усилению рефлекторной возбудимости и возникновению судорог, особенно глотательных и дыхательных мышц, к увеличению слюно- и потоотделения.
Иммунитет. Люди и животные, вакцинированные классическими вакцинами против бешенства, отвечают подъемом уровня нейтрализующих антител. Степень подъема определяется силой вакцины. Антитела направлены к G-гликопротеину (гемагглютинину). Сывороточные антитела у людей, больных бешенством, редко появляются до 8 дня болезни. В спинномозговой жидкости антитела появляются на 9-11-й дни. Клеточный иммунитет при бешенстве изучен недостаточно. Остается непонятным тот факт, что у 85 % людей, укушенных заведомо бешеным животным, бешенство не развивается, заболевают только 15 % пострадавших.
Клиника. При условии развития клинических признаков заболевания бешенство – абсолютно смертельная болезнь. Заболевание характеризуется неуклонным повышением температуры тела до 42˚С, развитием депрессии, апатии, расстройствами дыхания и глотания. При попытке питья и при виде воды возникают приступы гидрофобии – чувство ужаса и болезненные спазмы мышц глотки и гортани. Приступы гидрофобии могут быть спровоцированы движением воздуха (аэрофобия), ярким светом (фотофобия), громким звуком (акустофобия). Приступ сопровождается болезненными судорогами, агрессивностью, помрачением сознания. Через 1-2 дня от начала болезни появляется обильное слюно- и потоотделение. В последующем развиваются параличи нижних конечностей и черепно-мозговых нервов. Смерть наступает от паралича сердца или дыхательного центра. Общая продолжительность болезни – 3-7 дней.
Лабораторная диагностика. Используется вирусоскопический метод и биологическая проба на животных. При световой микроскопии тканей больших полушарий, мозжечка, продолговатого мозга и гиппокампа в цитоплазме нейронов обнаруживаются тельца Бабеша-Негри. Для их выявления используются способы окраски по Туревичу, Муромцеву-Селлеру и др. Тельца Бабеша-Негри имеют различную величину и форму (от 0,25 до 25 мкм, округлой или овальной формы). Тельца окрашиваются в красно-оранжевый цвет и выявляются в 90-95 % случаев в мозге собак, погибших от бешенства, и в 70 % случаев – у погибших от бешенства людей (см. главу 7). При отрицательных результатах патогистологических исследований применяют люминесцентную микроскопию, а также биологическую пробу на мышах. Для выявления антигенов вируса бешенства используют непрямой и прямой метод флюоресцирующих антител. Мазки-отпечатки ткани мозга фиксируют в ацетоне при температуре 4˚С в течение 8-10 ч, после чего на препараты наносят антирабический иммуноглобулин, меченный флюорохромами, и выдерживают во влажной камере при 37˚С в течение 30 минут. После этого препараты промывают буферным раствором (рН 7,4), сушат и исследуют в люминесцентном микроскопе. Нормальная ткань мозга флюоресцирует тусклым желтым цветом, антигены вируса бешенства – в виде зеленых гранул различной величины и формы. Биологическая проба проводится путем внутримозгового заражения под наркозом 15-20 мышей-сосунков или мышей в возрасте 3-5 дней. Инфицирующая доза – 0,03 мл суспензии исследуемого материала. Животные наблюдаются в течение 21 дня от момента заражения. Клинические проявления бешенства у мышей развиваются не ранее 5 дня. Идентификацию вирусов бешенства проводят в материале от погибших животных при использовании люминесцентной микроскопии.
Лечение. Не разработано. Проводится симптоматическое лечение.
Профилактика. Бешенство можно контролировать на 3 уровнях: человек, домашние животные и дикие животные. Имеются вакцины для профилактики среди людей и домашних животных (культуральные инактивированные вакцины, вводимые парентерально). Среди диких животных используется пероральная вакцина.
Предупреждение заболевания человека бешенством после укуса или ослюнения его бешеным или неизвестным животным осуществляется тщательной первичной обработкой раны с промыванием ее перекисью водорода и прижиганием концентрированной йодной настойкой. Затем проводят срочную серотерапию (антирабический γ-глобулин) и вакцинацию на пастеровских пунктах согласно инструкции, утвержденной Министерством здравоохранения Украины.
В настоящее время для профилактики бешенства используется антирабическая инактивированная культуральная сухая вакцина. Вакцина получена Селимовым (1974 г.) методом пассажей вирусов бешенства (штамм-fixe) в культуре клеток почек сирийского хомяка при 32˚С. Вакцинный штамм – Внуково-32 – инактивирован ультрафиолетовыми лучами. Вакцина выпускается в ампулах по 3 мл (после разведения). Вводится подкожно в область живота. В зависимости от категории повреждения и данных о животном разовая доза составляет от 2 до 5 мл. Основной курс вакцинации – от 7 до 21 дня, сроки проведения ревакцинации – согласно инструкции по применению вакцины. Вакцина используется для проведения экстренной профилактики бешенства (укусы, ослюнение бешеными животными). При вакцинации и после ее проведения (даже если она была прервана) употребление алкогольных напитков запрещено в течение 6 месяцев. Антитела появляются через 2 недели после начала прививок, при этом поствакцинальный иммунитет становится действенным примерно через 2 недели после окончания вакцинации. Вакцинация предупреждает возникновение бешенства в 96-99 % случаев. Иммунитет сохраняется в течение 1 года.
Неспецифическая профилактика бешенства городского типа предусматривает предупреждение бродяжничества собак и кошек, обязательную их регистрацию, профилактическую иммунизацию домашних животных, санитарно-ветеринарную пропаганду. Борьба с эпизоотиями природного типа имеет целью поддержание популяции животных, являющихся резервуаром бешенства, на определенном уровне.
Дата добавления: 2015-08-12 ; просмотров: 1894 . Нарушение авторских прав
источник
Перетрухина А. Т., Блинова Е. И.,
Возбудитель бешенства относится к роду Lyssavirus (от греч. lyssa — водобоязнь), семейству Rhabdoviridae.
Вирус бешенства единственный из царства Virae, который поражает всех теплокровных животных, в том числе человека, в глобальном масштабе с летальностью 100 %.
Род Lyssavirus объединяет 7 генотипов и 4 неклассифицированных вируса. 1-й генотип включает вирусы классического бешенства, обозначенные как штаммы уличного (дикого) вируса, выделенные от наземных млекопитающих, насекомоядных, плотоядных и кровососущих летучих мышей, а также фиксированные (вакцинные) штаммы вируса бешенства. Широко распространены в Европе, Азии, Северной и Южной Америке. 2-й генотип (Лагос-Бат) включает вирусы, выделенные от плотоядных летучих мышей, собак и кошек в Центральной и Южной Африке. 3-й генотип (Мокола) выделен от землероек, человека, собак и кошек в Центральной и Южной Африке. 4-й генотип (Дувенхаге) выделен от человека, укушенного летучей мышью, и летучих мышей в Южной Африке и Зимбабве. 5-й генотип (лиссавирус европейских летучих мышей 1-го типа) циркулирует в Европе, в т.ч. в европейской части России. 6-й генотип (лиссавирус европейских летучих мышей 2-го типа) циркулирует в Европе. Выделен также от человека в Финляндии и Шотландии. 7-й генотип выделен от людей в Австралии.
Вирион имеет форму пули размерами 170×70 нм и состоит из несегментированного генома, представленного одной молекулой спирально закрученной РНК негативной полярности и 5 структурных белков, из которых белок N является группоспецифическим антигеном.
РНК не инфекционна, а рибонуклеопротеид, в составе которого находится транскриптаза, обладает инфекционными свойствами.
В составе вириона, помимо РНК-полимеразы, содержится ряд ферментов. Среди них находится протеинкиназа, полиаденилаттрансфераза и другие ферменты.
Для вируса бешенства характерна адаптационная изменчивость, которая была использована Л. Пастером для получения вакцины против бешенства. Уличный вирус бешенства пассировался интрацеребрально на кроликах. По мере пассирования длительный инкубационный период, характерный для инфекции, вызванной уличным вирусом, постепенно сокращался, пока не стал равным 5 дням. Этот вирус был назван фиксированным и при интрацеребральном введении он вызывал 100 % гибель кроликов. Вместе с тем, он потерял патогенные свойства для собаки и человека. Поэтому фиксированный вирус, после дополнительной обработки (высушивание мозга кролика, приводящее к инактивации вируса) был использован в качестве вакцины для лечебно-профилактических прививок.
Бешенство — острое нейровирусное заболевание, передаваемое человеку через укус лисы, собаки, волка и других плотоядных животных, зараженных в природе, в свою очередь, так же — через укус.
После укуса больного животного вирус размножается в мышечной ткани в месте укуса, а затем, достигнув нервных окончаний чувствительных периферических нервов, распространяется центростремительно, достигая двигательных нейронов. Происходит распространение вируса в нервно-мозговой ткани, демиелиницация белого вещества, дегенерация аксонов и миелиновых оболочек. В спинном мозге более всего поражаются задние рога. В цитоплазме пораженных нервных клеток образуются специфические цитоплазматические включения — тельца Бабеши-Негри, которые являются диагностическим признаком болезни. Включения ацидофильны, четко контурированы, имеют сферическую форму с диаметром от 0,25 до 25 мкм, чаще 2-10 мкм. Размножение вируса происходит в слюнных железах. К моменту появления первых клинических признаков бешенства вирус широко диссеминирован во всем организме. В связи с этими особенностями патогенеза инкубационный период при бешенстве продолжителен, и тем продолжительнее, чем дальше от спинного и головного мозга находится место укуса. Под инкубационным периодом подразумевается период времени от момента укуса до появления первых признаков болезни. Он варьирует от 6-7 суток до 6 лет, но описаны примеры инкубационного периода продолжительностью 10, 11, 13 и 19 лет. Средний инкубационный период при бешенстве составляет 1-3 месяца. Таким образом, длительность его зависит от продолжительности репликации вируса в воротах инфекции, места и тяжести укусов, количества попавшегов рану вируса, свойств вируса, количества нервных окончаний на месте укуса, а также структуры популяции штамма вируса.
Заболевание у человека начинается с продромального периода, который длится 2-4 дня и проявляется недомоганием, головной болью, тошнотой, рвотой, повышением температуры. Во входных воротах инфекции нарушается чувствительность. Повышается активность симпатической нервной системы: отмечаются слезотечение, расширение зрачков, потливость, обильное слюноотделение, при глотании — болезненные спазмы. У больного появляется чувство страха, особенно при виде воды. Появляются агрессивность и буйство, конвульсивные судороги, в финальной стадии — параличи, упадок сердечной деятельности и наступает смерть, обычно через 3-5 дней после начала заболевания.
Обычно заболевание имеет летальный исход и поэтому иммунитет после перенесенной инфекции не изучен. После вакцинации людей убитой антирабической вакциной появляются антитела, сохраняющиеся в течение года. Ревакцинация приводит к резкому повышению титра антител.
Лабораторная диагностика включает обнаружение телец
Бабеши-Негри в тканях мозга (посмертная) и выделение вируса молекулярно-генетическими методами ОТ-ПЦР и секвенированием генов N- и G-белков, а также выявление рибонуклеопротеина прямым методом флюоресцирующих моноклональных антител (прижизненная).
Бешенство у человека неизлечимо и всегда заканчивается летальным исходом. Единственный метод борьбы с бешенством —
профилактика. Профилактика состоит из предупреждения инфицирования человека и предупреждения развития бешенства у инфицированного человека. Предупреждение инфицирования включает контроль за численностью основных носителей вируса в природе и бездомных и домашних животных, иммунизацию домашних животных от вируса бешенства. Предупреждение развития бешенства у человека после контакта с бешеным или подозрительным на бешенство животным осуществляется путем оказания немедленной антирабической помощи. Антирабическая помощь состоит из местной обработки раны, введения антирабической вакцины или комплексного применения антирабического иммуноглобулина
и вакцины.
Для лечебно-профилактических прививок в РФ и развитых странах применяются концентрированные, очищенные, высокоиммуногенные культуральные вакцины, приготовленные на различных клеточных субстратах, а также антирабический иммуноглобулин. В развитых странах используется человеческий антирабический иммуноглобулин, а в РФ и третьих странах — лошадиный антирабический иммуноглобулин. При поверхностных укусах и царапинах применяется одна вакцина. В РФ вакцина вводится шестикратно внутримышечно по 1 мл в 0, 3, 7,14, 30, и 90-е сутки. В развитых странах вакцина применяется 5-кратно.
Вакцина антирабическая культуральная концентрированная очищенная инактивированная сухая (Рабивак — «Внуково-32»), лиофилизат для приготовления раствора для внутримышечного введения и антирабического иммуноглобулина (КОКАВ)
Препарат применяется для лечебно-профилактической и профилактической иммунизации человека против бешенства. Вакцина антирабическая культуральная концентрированная очищенная инактивированная сухая, лиофилизат для приготовления раствора для внутримышечного введения представляет собой вакцинный вирус бешенства штамм «Внуково-32», выращенный в первичной культуре клеток почек сирийских хомячков, инактивированный ультрафиолетовыми лучами и формалином, концентрированный методом ультрафильтрации с последующей очисткой методом гельхроматографии.
Вакцина антирабическая — пористая масса белого цвета, гигроскопична.
Одна доза (1,0 мл) вакцины содержит: специфический антиген вируса бешенства штамм «Внуково-32» — не менее 2,5 ME (Международных Единиц) — активный компонент, альбумин — 5,0 мг (раствор для инфузий) — стабилизатор, сахарозу — 75,0 мг — стабилизатор, желатин — 10,0 мг — формообразователь. Препарат не содержит консервантов и антибиотиков.
Вакцина выпускается в комплекте: 1 ампула вакцины по 1,0 мл (1 доза) и 1 ампула растворителя (вода для инъекций) по 1,0 мл. В упаковке содержится 5 комплектов (5 ампул с вакциной и 5 ампул с растворителем).
Вакцина индуцирует выработку иммунитета против бешенства. Вакцинацию проводят по следующей схеме: первичная иммуниза-
ция — 3 инъекции в 0, 7 и 30 день по 1,0 мл; первая ревакцинация через 1 год — 1 инъекция 1,0 мл; последующие ревакцинации через каждые 3 года — по 1 инъекции 1,0 мл.
Вакцину хранят и транспортируют при температуре от 2 до 8 °С. Допускается транспортирование вакцины при температуре до 25 °С в течение не более 2-х суток.
Срок годности вакцины — 2 года.
Сухая антирабическая вакцина типа Ферми
Вакцину изготовляют из мозга овец в возрасте до 1 года, зараженных фиксированным вирусом бешенства.
Для приготовления вакцины по методу Ферми измельченный мозг разводится физиологическим раствором, содержащим 1 % фенола, так, чтобы получить 5 % взвесь. После этого вакцина для инактивации выдерживается в течение 8 дней при 20-22 °С. После инактивации вакцина фильтруется через четырехслойный марлевый фильтр и после проверки стерильности передается в разливку.
Вакцину выпускают в сухом виде. Она представляет собой таблетку беловато-серого цвета. Одна таблетка получается в результате лиофильного высушивания 0,75 мл 10 % вируссодержащей суспензии. В коробку с 5 ампулами сухой вакцины вкладывают 5 ампул, содержащих по 3 мл растворителя — дистиллированной воды. После растворения вакцина представляет собой гомогенную жидкость беловато-серого цвета. Вакцину вводят немедленно после разведения. Хранение разведенной вакцины не допускается.
Срок годности сухой вакцины — 3 года.
Рабипур — инактивированная очищенная вакцина против бешенства
Рабипур — вакцина для постэкспозиционной (после укуса бешеного или подозрительного животного) и профилактической вакцинации против бешенства.
В настоящее время вакцина находится на перерегистации.
Рабипур получают путем выращивания штамма вируса Flury LEP на культуре куриных фибробластов. 1 доза лиофилизированного порошка содержит инактивированный вирус бешенства (штамм Flury LEP) с активностью более 2,5 МЕ. Выпускается во флаконах по 1 дозе, в комплекте с иглой и растворителем в ампулах по 1 мл. Вакцина не содержит консервантов.
Порошок белого цвета, после растворения — бесцветная жидкость.
Хранят при температуре 2-8 °C.
Срок годности — 3 года.
Жидкая антирабическая вакцина МИВП
Вакцина представляет собой 5 % суспензию в 0,96 % растворе фенола головного мозга белых крыс-сосунков, зараженных фиксированным вирусом бешенства, в возрасте 4-8 дней. Этот препарат выпускается в жидком виде.
Срок годности жидкой вакцины — 5 месяцев.
Сухая антирабическая вакцина МИВП
Для приготовления сухой вакцины к 20 % суспензии головного мозга белых крыс-сосунков, инактивированной в фенолизированном физиологическом растворе при 22 °С в течение 14 дней, добавляют равный объем стабилизирующей сахарозо-желатиновой среды, разливают в ампулы по 3 мл и высушивают из замороженного состояния.
В коробку с 10 ампулами сухой вакцины помещают равное количество ампул, содержащих по 3 мл дистиллированной воды, применяемой в качестве растворителя.
Срок годности сухой вакцины — 1 год.
Культуральная антирабическая вакцина
Препарат представляет собой вируссодержащую жидкость, полученную при выращивании фиксированного вируса бешенства на первичной культуре клеток почки сирийских хомяков и частично инактивированную действием фенола. Для приготовления вакцины используется штамм «Сад», идентичный с фиксированным вирусом. Он аттенуирован интрацеребральными пассажами на белых мышах и выращиванием в культуре тканей.
Срок годности вакцины — 1 год.
Антирабический иммуноглобулин из сыворотки лошади
Препарат изготавливается из сыворотки лошадей, гипериммунизированных вирусом бешенства. Выпускается в жидком виде в ампулах и флаконах по 5 и 10 мл. На ампулах и коробках с антирабическим гамма-глобулином, кроме других необходимых данных, указывается титр в международных единицах (МЕ). Активность должна быть не менее 800 МЕ/мл, в коробки вкладывают также ампулы с разведенным в 100 раз препаратом, который применяют для определения чувствительности к лошадиному белку.
По условным показаниям антирабический иммуноглобулин вводят взрослым в дозе 0,25 мл/кг, по безусловным — 0,25-0,5 мл/кг. Всю дозу иммуноглобулина вводят в один день.
Антирабический иммуноглобулин должен храниться при 2-8 °С. Срок хранения — 2 года.
Иммуноглобулин антирабический из сыворотки крови человека
Применяют в комбинации с антирабической вакциной для предупреждения заболевания людей гидрофобией при тяжелых множественных укусах бешеными или подозрительными на бешенство животными.
При повторном множественном укусе больным бешенством или подозрительным на бешенство животным антирабический иммуноглобулин не назначается, если при первом укусе пострадавший получил полный комбинированный курс лечения антирабическим иммуноглобулином и антирабической вакциной.
Препарат представляет собой концентрированный раствор очищенной гамма-глобулиновой фракции сыворотки крови человека, выделенной методом холодовой экстракции этанолом и подвергнутой процессу ультрафильтрации, очистки и вирусной инактивации при значении рН 4,0 и температуре 23-25 °С в течение
21 дня.
В 1 мл содержит специфические антитела к вирусу бешенства, не менее 150 ME; стабилизатор глицин (гликокол) от 20 до 25 мг; натрия хлорид 7 мг; вода для инъекций. Препарат не содержит антибиотиков. HBsAg, антитела к ВИЧ-1, ВИЧ-2 и к вирусу гепатита С отсутствуют. Прозрачная или слегка опалесцирующая жидкость, бесцветная или светло-желтого цвета.
Препарат вводят в как можно более ранние сроки после обращения однократно в дозе 20 МЕ/кг массы тела взрослого или ребенка. Введение антирабического иммуноглобулина в более поздние сроки, а также, после введения антирабической вакцины не допускается. Дозировка иммуноглобулина не должна превышаться ни при каких обстоятельствах, так как введение повышенной дозы иммуноглобулина может частично подавить продукцию
антител.
Хранение при температуре от 2 до 8 °С в защищенном от света месте. Не допускается замораживание.
Срок годности препарата — 2 года.
Гомологичный антирабический иммуноглобулин из сыворотки людей, иммунизированных против бешенства
Препарат представляет собой прозрачную или слегка опалесцирующую жидкость, бесцветную или имеющую легкую желтую окраску. В 1 мл препарата должно содержаться не менее 40 МЕ антител против бешенства. Доза препарата не менее 160 МЕ/кг.
Срок годности иммуноглобулина — 1 год.
Флюоресцирующий антирабический гамма-глобулин
Для ускоренной индикации антигена вируса бешенства прямым иммунофлюоресцентным методом применяется антирабический гамма-глобулин, химически связанный с флюоресцеинизотиоцианатом. Он сообщает антигену вируса бешенства, фиксированного в отпечатках мозга, яркую специфическую флюоресценцию при разведении конъюгата не ниже 1:8. В отпечатках мозга, содержащего другие гетерологичные вирусы, флюоресценция отсутствует.
Срок годности препарата — 1 год.
источник
Бешенство – особо опасное инфекционное заболевание, вызываемое инфицированием нейтропным вирусом Rabies virus . Провоцирует возникновение энцефалита (воспалительного процесса головного мозга) специфического происхождения. При попадании в живой организм вирус распространяется по клеткам ЦНС, инициирует разрушение их структурной целостности с последующим отмиранием. Патология характеризуется быстрым течением активной фазы, тяжелой симптоматикой и высоким риском летального исхода.
Заражение человека происходит вследствие укуса инфицированного животного. Эволюционная частотность болезни зависит от места инфицирования и тяжести локального поражения. Само заболевание характерно специфическими признаками на всех стадиях развития.
Этиология вируса доподлинно не изучена. Существуют рукописные доказательства происхождения заболевания еще с древних времен. Тогда болезнь носила название «водобоязнь» (гидрофобия) – древние люди замечали у зараженных животных паническую боязнь воды. В то время эффективных лекарств против бешенства не придумали, поэтому укушенный человек заранее обрекался на смерть.
Вирус бешенства классифицирован по миксовирусовой группе Lyssavirus семейство Rhabdovtridae. Имеет форму винтовочной пули размерами до 200 нм с однонитиевой РНК. Является патогенным для теплокровных животных и человека. Различают два вида вириона: циркулирующий (природный, опасный для людей) и фиксированный (синтезирован лабораторно, безвредный для человека).
Источником заражения человека могут быть собаки (до 60%), кошки, дикие животные (до 35%), волки (3%), другие животные (2%). Существует гипотеза первоначального источника вириона бешенства – грызуны, не погибшие сразу после вирусного проникновения.
Вирион бешенства концентрируется в слюнных железах, выделение активного микроба происходит со слюной жидкостью. Вероятность заражения человека больным животным при укусе составляет 95%, при ослюнении 60%. Вирион-возбудитель бешенства обладает высокой тропностью (органическая привязанность к определенному виду ткани) к нервным клеткам.
Первоначально микроб активизирует деятельность в мышечных волокнах раны, затем перемещается к нервным окончаниям. Посредством деления и размножения бактерия разрушает нервную систему, образуя на месте погибших клеток тельца Бабеша-Негри. Органические образования представляют собой специфические включения цитоплазмы в репликационных местах.
Судороги и параличи инфицированного человека обусловлены вирусным достижением нервной системы головного мозга с последующим стремительным разрушением жизнеобеспечивающих участков мозгового вещества. На этой стадии излечение пациента невозможно, больной впадает в коматозное состояние и умирает.
- микроб устойчив к низкому перепаду температуры, в замороженном состоянии сохраняет активную жизнедеятельность;
- в трупах животных содержится до 4-х месяцев;
- против бешенства бессильны любые сильнодействующие антибиотики, антибактериальные препараты, противовирусные средства;
- продвигаясь по нервным клеткам, вызывает воспалительные процессы, разрушающие их структуру.
Развитие и прогрессирование бешенства после укуса напрямую связано с поздним сроком обращения укушенного за медицинской помощью, нарушения прививочного режима или комплекса курса терапии. Наибольшая распространенность возбудителя болезни обуславливается нахождением человека в сельской местности вблизи лесных насаждений.
Период инкубации бешенства длиться от 10 дней до 4-х месяцев, чаще всего срок сокращается до 1 месяца. На практике зафиксированы случаи более долгого инкубационного периода – от года до 5 лет.
Вероятность прогрессирования болезни зависит от воздействующих факторов: вид инфицированного животного, количество попавших в рану вирионов, сопротивляемости иммунной системы, место заражения (наиболее восприимчивыми являются участки тела с скоплением нервных окончаний).
Специфические симптомы развития болезни условно подразделяются на несколько этапов, различимых между собой характерными признаками и поведением больного.
- Ранний (продромальный). Длительность до трех дней. Наблюдается субфебрильная температура, отсутствие или нарушение сна, апатия, угнетенность, беспокойное поведение пациента. В месте заражения ощущаются болезненные симптомы, даже если укус зарубцевался.
- Разгар (агрессия). Длится до 4 дней. Резкая гиперчувствительность к любому внешнему раздражителю – яркое освещение, громкий звук и шум. Пациент боится воды, агрессивен, испытывает дикое чувство страха, судорожно вздрагивает. В этот период появляются галлюцинации и бредовые видения, неадекватное поведение и реакции больного, повышенное слюноотделение. Иногда пациент замирает, начинает тяжело дышать и смотреть в одну точку расширенными зрачками глаз.
- Параличный (бешенство). Вследствие паралича скуловой и глазной мускулатуры отвисает нижняя часть челюсти. Поздняя стадия характеризуется неестественным аппетитом (больной есть несъедобные и опасные вещи). Наблюдается полная деградация пациента как осознанной личности. По причине паралича дыхательных путей наступает удушье.
Совокупная продолжительность активной фазы болезни составляет от 8 до 12 дней. Однако не исключаются атипичное прогрессирование заболевания – отсутствие продромальной стадии или начало болезни с параличного периода. В большинстве атипичных случаев диагностировать болезнь удается только посмертно.
Диагностика симптомов бешенства проводится путем сбора и тщательного анализа данных анамнеза заболевания у пострадавшего человека.
- Фундаментальное значение в длительности временного промежутка между инкубационным периодом и проявлением видимых признаков имеет место заражения. Больной осматривается на предмет характерных особенностей раны, глубины проникновения зубов животного, степени ослюнения.
- Пациент визуально диагностируется врачом по первоначальным признакам болезни – расширены ли зрачки, как ведет себя больной, что чувствует.
- Для микрообследования производят забор клеток слюнной железы и заражают ими лабораторных мышей. Наличие бешенства определяется по быстро развивающимся признакам паралича и смерти грызунов.
- Так как специфическое исследование вируса бешенства затруднительно выполнить при жизни больного в острой стадии течения заболевания, окончательный диагноз устанавливается после смерти пациента.
Появлению видимых симптомов болезни у домашних животных предшествует возникновение вирусного возбудителя в слюне.
Любое домашнее животное становиться заразным примерно за неделю до появления характерных признаков бешенства и остается опасным весь период острого течения болезни.
Заражение собаки происходит от укуса других собак, волоков или кошек. Стандартный период инкубации у собаки от 3 до 7 дней.
Продромальная стадия начинается с беспокойного поведения пса. У животного появляется несвойственный аппетит – поедание абсолютно несъедобных предметов. Лай воющий с сиплым оттенком. Собака старается грызть укус, появляются признаки возбуждения или чрезмерной ласковости, характерно слюнотечение.
Фаза возбуждения начинается с 2-3 дня болезни. Сопровождается странным поведением животного – вспышки агрессии резко сменяются нападением, пес грызет землю, периодически появляются судорожные сокращения мышц, рвота и косоглазие.
Появление стадии депрессии (параличной) определяется по признакам: собака истощена, ослаблена. На 4 день наступает паралич и гибель животного.
Заражение пушистого домашнего питомца происходит от укусов больных грызунов, собак, ежей и других котов. Латентный период инкубации длится от 10 дней до 3 недель, у котят до 7 дней максимум.
Симптомы бешенства у кошек развиваются на протяжении 3-10 дней. В начальном периоде животное вялое, не ест, резко сильно ласковое. Далее начинает развиваться пугливость, беспокойное поведение и беспричинная нервозность. Кошка кусает и грызет несъедобные вещи, озирается по сторонам, пытается царапать место укуса. Не исключаются появление тошноты, рвоты, жидкого стула. Приступ агрессии может вызвать громкий шум, стук, яркий свет.
На 3-е сутки развивается маниакальная стадия и длится до 5 дней. Обильное слюнотечение сменяется спазмами мышц глотки. Кошка не может самостоятельно пить, старается напасть и укусить.
Срок депрессивной стадии не более 2 суток. Животное впадает в апатию, не двигается. Кошка не может мяукать, отвисает нижняя челюсть, выпадает язык, мутнеет роговица глаз. Наступает онемение конечностей и смерть животного от парализации дыхательных мышц.
Многие люди задумываются о том, а стоит ли вообще делать прививку от бешенства ? На практике давно существует бесплатная ветеринарная услуга – вакцинация животных от бешенства. Ежегодное применение вакцины от бешенства в профилактических целях – это залог вашего здоровья и жизни домашнего любимца.
Фундаментальные правила профилактических мер заключаются в годовой поголовной вакцинации домашних животных, введение карантина при обнаружении очага инфекции.
Единственная возможность спастись от смерти – предотвратить появление визуальных проявлений болезни. Вакцину от бешенства необходимо ввести до 14 дня с момента укуса животного. Наиболее рекомендуемая форма вакцинации – укол специфического иммуноглобулина или активная периодичная иммунизация, после которой категорически запрещается употребление алкоголя на срок до полугода.
С момента заражения через укус современные вакцины используются не более 6 раз: в первый день обращения к врачу, затем на 3, 7, 14, 30 и 90 день соответственно. Укусившее животное изолируется, за ним устанавливается медицинское наблюдение в течение 10 дней. Если поведение животного не вызывает подозрений, вакцинация потерпевшего прекращается.
Зарегистрированные очаги бешенства России природной этиологии
- Поволжье (западные и центральные районы). В 72% случаях источником заражения являются лисицы. Распространяется волками, енотами и барсуками.
- Заполярье (арктическая циркуляция). Вирус распространяется среди песцов.
- Городские очаги поражения. Вирус популяризуется среди собак, источников заражения чаще всего становятся кошки и сельскохозяйственные домашние животные.
Поражение бешенством человека – это как игра со смертью, не сегодня и не завтра. Но не повезет. Если вас укусило животное, нельзя думать что все заживет и будет хорошо. Помните, незамедлительное обращение за медицинской помощью до истечения 14 дней после укуса – 99% вероятность выживания даже при условии инфицирования вирионом бешенства.
источник
Бешенство(rabies)- острая инфекция из группы зоонозов, вызываемая вирусами из семейства рабдовирусов и протекающее с тяжелым поражением нервной системы и заканчивающееся, как правило, смертельным исходом. История Еще Аристотель обращал внимание на распространение бешенства среди собак. Цельс в I в. н.э. указывал на частые заболевания собак и других животных. Он заметил и другие факты: заболевания бешенством людей, возможность передачи болезни со слюной животного в момент укуса. В 1271 году описана крупная эпизоотия бешенства среди собак и волков в Германии, в 1500 году в Бельгии и Испании. В 1590 году Отмечена эпизоотия бешенства среди волков. Изучение бешенства в 19 веке тесно связано с достижениями Пастера, Мечникова, Гамалеи. В 1885 г. Л. Пастер получил и с успехом использовал вакцину для спасения людей, укушенных бешеными животными. Вирусная природа болезни доказана в 1903 г. П. Ремленже, который произвел фильтрование заразного материала через бактериальные фильтры. XIX Таксономия Возбудитель бешенства РНК-содержащий вирус Neuroryctes rab /> Вирион бешенства в препаратах инфицированного мозга и культур ткани под электронным микроскопом имеет пулевидную форму с одним плоским и одним закругленным концом. Имеет спиральный нуклеокапсид, окруженный белково-липидной оболочкой с поверхностными выступами. Диаметр вириона 75-80 нм, длина 180 нм. Геном – однонитчатая РНК с молекулярной массой 3,5-4,6 106. Вирионы содержат 5 структурных белков и РНК-зависимую полимеразу. В поперечных срезах вириона бешенства виден центральный стержень, окруженный плотной оболочкой. Для морфогенеза вируса бешенства характерно первоначальное формирование матрикса; филаментозной субстании в цитоплазме инфицированных клеток. В последующем из матрикса образуется нуклеокапсид вириона. Виды бешенства Существование вируса бешенства за счет различных животных и на различных территориях привело к тому, что он оказался неоднороден: у отдельных представителей имеются различия по вирулентности, по характеру вызываемого инфекционного процесса и патологоанатомических изменений. В антигенном отношении популяция вируса более или менее однотипна, хотя некоторые различия у разных вирусов теперь обнаружены. В частности, оказалось, что пастеровский вакцинный фиксированный штамм (PV-11) по антигенной структуре полноценнее, универсальнее многих выделяемых сейчас от человека и животных. Различают следующие варианты вируса бешенства: Типичный (уличный) вирус бешенства, в популяции которого обнаружены в свою очередь определенные различия. В частности, лисьи штаммы отличаются по патогенности. Патологоанатомические изменения в мозгу при бешенстве, которое вызывается укусом лисицы, отличаются тем, что тельца Бабеша—Негри встречаются существенно реже, изменения в ЦНС носят более диффузный характер в виде очень мелких бесструктурных плазматических включений. Вирус африканского собачьего бешенства. Этот вирус слабопатогенен для лабораторных животных. Заболевания людей, вызванные этим вирусом, или не встречаются вовсе, или они чрезвычайно редки. Вирус дикования — болезни собак и песцов в северных тундровых районах. Человек болеет очень редко — описано заболевание одной девочки на Аляске. Имеются наблюдения о развитии в результате лабораторного заражения медленной нейроинфекции, закончившейся смертью. Вирус американского бешенства летучих мышей — наиболее существенно отличающийся от основного вируса типичного бешенства. Среди вирусов, выделяемых от летучих мышей, встречается несколько разновидностей. По современным представлениям все названные вирусы относятся к одному виду — вирусу Neuroryetes. Известен еще один вариант вируса бешенства — фиксированный, полученный Пастером при адаптации уличного вируса в организме кролика в измененных условиях заражения. Вирус бешенства патогенен для человека и всех видов теплокровных животных, а также птиц, но степень восприимчивости к вирусу бешенства у разных видов животных различна. Согласно экспериментальным данным наибольшей чувствительностью к вирусу бешенства обладают лисицы, среди лабораторных животных – сирийский хомяк.Фиксированный вирус не выделяется со слюной и не может быть передан во время укуса. Механизм вирусной персистенции в клеточных культурах связывается с образованием и накоплением Ди-частиц. Проникновение вируса в клетки происходит путем адсорбционного эндоцитоза — вирионы выявляются в виде включений, окруженных мембраной, адсорбированных на микротрубочках и в составе лизосом. Нейрон, инфицированный вирусом бешенства. Видны вирусы в цитоплазме нейрона Штаммы вируса бешенства циркулирующие в различных географических зонах мира, обладают рядом общих биологических и антигенных характеристик (вариабельность инкубационного периода, поражения пи переферическом введении ЦНС, выделением вируса со слюной, тождественной антигенной структурой.) Наряду с этим установлено существование естественных биовариантов вируса бешенства. Биоварианты вируса бешенства различаются по степени патогенности при внемозговом заражении, распределению в органах зараженных животных, способности формировать включения. Вирусы подобные возбудителю бешенства, имеют частичное антигенное с ним родство, а также сходный тип репродукции, но отличаются по комплексу биологических характеристик. Химический состав Очистка и концентрация культурального вируса позволили изучить химическую структуру и биологическую активность субвирусных компонентов. В состав вириона бешенства входят: РНК(1%), белки(72%), липиды(24%), углеводы(1%). Белки вириона бешенства представлены 4 высокомолекулярными и 1 низкомолекулярным полипептидами. Нуклеокапсид вириона содержит 96% белка, представляет собой однонитчатую правовращающуюся спираль РНК длиной 1 мкм и наружным диаметром 15-16 нм, окруженную оболочкой. Состоящей из гликопротеина и глюколипидов. Нуклеокапсид вириона бешенства обладает константой седиминтации 200S, плавучей плотностью 1,32 г/см 3 , молекулярным весом 1,5 Ч 10 8 дальтонов и константу седиментации 45S. Свойства Вирус бешенства обладает антигенным, имунногенными и гемагглютинирующим свойствами, однако биологическая и антигенная активность различных субвирусных структур неодинакова. Белки оболочки обладают выраженной иммуногенной активностью, в то время как белки «ядра» вириона мало иммуногены. Гликопротеин оболочки вириона обладает гемагглютинирующей активностью. Очищенная вирусная РНК, нуклеокапсид и вирусные компоненты «ядра» вириона не инфекциозны, что может быть связано с отсутствием в их составе гликопротеина оболочки, необходимого для адсорбции вируса на поверхности клетки. В составе вириона бешенства обнаружены собственные ферментные системы: протеинкиназа, ДНК-полимераза. Синтез вириона происходит как в веществе матрикса, так и в окружающих его интрацитоплазматических мембранах. В цитоплазме нейронов зрелые вирионы освобождаются путем почкования от поверхности филаментозного матрикса. В культуре клеток зрелые вирионы отпочковываются от клеточной поверхности. Размножение вируса вызывает в аммоновом роге головного мозга и среднем мозге поражение сосудов с образованием в цитоплазме нейронов специфических включений – телец Бабеша-Негри. Выделяется со слюной. Размножение вируса бешенства in vivo и in vitro, сопровождается фрмированием специфических включений – телец Бабеша-Негри. Тельца Бабеша-Негри размером 0,5-25 мкм расположены в цитоплазме нейронов, имеют округлую, овальную, реже веретенообразную форму, базофильную внутреннюю структуру, окрашивающуюся кислыми красителями в рубиновый цвет. При цитохимическом исследовании телец установлено, что они содержат РНК. Электронномикроскопическое исследование телец выявило, что их эозинофильная основа соответствует специфическому филаментозному матриксу инфицированных клеток, а внутренние базофильные структуры состоят из вирусных нуклеокапсидов, связанных с компонентами клеток. Степень формирования телец Бабеша-Негри при заражении различных штаммов вируса бешенства определяется численностью локальных очагов вирусного синтеза в цитоплазме инфицированных клеток . Резистентность Устойчивость вируса бешенства невелика. Некоторые детергенты вызывают распад вирионов. Вирус устойчив к фенолу, антибиотикам. Разрушается кислотами, щелочами, нагреванием (при 56°С инактивируется в течение 15 мин, при кипячении — за 2 мин. Чувствителен к ультрафиолетовым и прямым солнечным лучам, к этанолу к высушиванию. Быстро инактивируется сулемой (1:1000), лизолом (1-2%), карболовой кислотой (3-5%), хлорамином (2-3%). Вирус хорошо сохраняется лишь при низкой температуре и после вакуумного высушивания и в замороженном состоянии. Эпидемиология Бешенство типичный зооноз, от человека к человеку вирус передается крайне редко. Источником заражения являются инфицированные животные (лисы, волки, собаки, кошки, летучие мыши, грызуны, лошади, мелкий и крупный рогатый скот). Заражение человека происходит при укусе или ослюнении животным поврежденной кожи или слизистой оболочки. Вирус выделяется во внешнюю среду со слюной инфицированного животного или человека. Описаны случаи заболевания людей в результате укусов внешне здоровым животным, продолжающим оставаться таковым в течение длительного времени. В последние годы доказано, что помимо контактного возможны аэрогенный, алиментарный и трансплацентарный пути передачи вируса. Не исключается передача вируса от человека к человеку. Описаны несколько случаев заражения людей в результате операции по пересадке роговой оболочки глаза.Различают эпизоотии бешенства природного и городского типов. Естественными являются природные эпизоотии бешенства, поддерживаемые дикими плотоядными животными из семейства собачьих, кошачьих и рукокрылых. В городских очагах источником бешенства являются домашние животные. Заразными считаются животные за 3-10 дней до появления признаков болезни и далее в течение всего периода заболевания. Часто больных бешенством животных можно отличить по обильному слюно-и слезотечению, а также при наблюдении признаков водобоязни. Заражение человека происходит при укусе «бешеным» животным Анализ заболевания показывает, что 53,5% заболевших бешенством в нашей стране вовремя не обратились за медпомощью и не получили антирабические прививки. Локализация укуса имеет решающее значение в частоте заболеваемости. Среди заболевших 67% были укушены в верхние конечности, 12% в нижние, 1,9% в туловище. В настоящее время вызывает опасения серьезный рост заболеваемости бешенством среди животных и людей. возрастает количество безнадзорных собак и кошек и неудовлетворительно осуществляется их отлов и изоляция. До сих пор не приняты правовые акты, регламентирующие правила содержания домашних животных. Только за 2009 число тех, кто пострадал от животных с диагнозом бешенство, составило более шести тысяч человек. Патологическая анатомия У людей погибших от бешенства, выявляется отек и набухание головного и спинного мозга с выраженной гиперемией, сопровождающейся петихиальными кровоизливаниями. Однако воспалительные процессы в тканях мозга часто являются ограниченными. Микроскопически воспалительный процесс обнаруживают в тройничных нервах, гассеровых узлах, межпозвоночных и верхних шейных симпатических ганглиях и в сером веществе головного мозга. В периферических нервах отмечают лимфоидные и моноцитарные инфильтраты, локализующиеся около расширенных сосудов. Вокруг гибнущих или распавшихся ганглиозных клеток возникает гиперплазия олигодендроглиальных элементов, в результате чего образуются так называемые узелки бешенства. Они, однако, не являются специфичными для бешенства, так как для бешенства так как встречаются и при других заболеваниях(энцефалит) Характерными для бешенства считают появление цитоплазматических включений-телец Бабеша-Негри, которые чаще всего обнаруживаются в клетках гипокампа и в грушевидных нейроцитах мозжечка. Во внутренних органах особых изменений не возникает. Слюнные железы иногда увеличиваются. В них обнаруживаются периваскулярные инфильтраты. Отмечается полнокровие и некоторая раздутость легких. Селезенка не увеличена. Патогенез Вирус бешенства попадает в организм человека при укусе либо ослюнении бешеным животным через рану или микроповреждения кожи, реже слизистой оболочки. Далее вирус бешенства распространяется по нервным стволам центростремительно, достигает центральной нервной системы, а затем опять-таки по ходу нервных стволов центробежно направляется на периферию, поражая практически всю нервную систему. Таким же периневральным путем вирус попадает в слюнные железы, выделяясь со слюной больного. Нейрогенное распространение вируса доказывается опытами с перевязкой нервных стволов, которая предупреждает развитие болезни. Тем же методом доказывается центробежное распространение вируса во второй фазе болезни. Скорость распространения вируса по нервным стволам составляет около 3 мм/ч. Одна из гипотез объясняет распространение вируса бешенства по аксоплазме периферических нервов к ЦНС влиянием электромагнитного поля организма на отрицательно заряженные вирионы [Аnnаl N., 1984]. В опытах на мышах удается достичь лечебного эффекта, подвергая животных воздействию электрического поля, создающегося путем фиксации отрицательного электрода на голове, а положительного — на лапке. При обратном расположении электродов наблюдается стимуляция инфекции. Нельзя отрицать также роль гематогенного и лимфогенного пути распространения вируса в организме. Интересно, что последовательность аминокислот гликопротеида вируса бешенства аналогична с нейротоксином змеиного яда, избирательно связывающимся с ацетилхолиновыми рецепторами. Возможно, этим обусловливается нейтротропность вируса бешенства, а связыванием его со специфическими нейротрансмиттерными рецепторами или другими молекулами нейронов объясняется развитие аутоиммунных реакций и селективное поражение некоторых групп нейронов. Размножаясь в нервной ткани (головной и спинной мозг, симпатические ганглии, нервные узлы надпочечников и слюнных желез), вирус вызывает в ней характерные изменения (отек, кровоизлияния, дегенеративные и некротические изменения нервных клеток). Разрушение нейронов наблюдается в коре большого мозга и мозжечка, в зрительном бугре, подбугорной области, в черном веществе, ядрах черепных нервов, в среднем мозге, базальных ганглиях и в мосту мозга. Однако максимальные изменения имеются в продолговатом мозге, особенно в области дна IV желудочка. Вокруг участков пораженных клеток появляются лимфоцитарные инфильтраты (рабические узелки). В цитоплазме клеток пораженного мозга (чаще в нейронах аммонова рога) образуются оксифильные включения (тельца Бабеша-Негри), представляющие собой места продукции и накопления вирионов бешенства. Клиническая картина Инкубационный период (период от укуса до начала заболевания) в среднем составляет 30-50 дней, хотя может длится 10-90 дней, в редких случаях — более 1 года. Причем чем дальше место укуса от головы, тем больше инкубационный период. Во время инкубационного периода самочувствие у укушенных не страдает. Особую опасность представляют собой укусы в голову и руки. Дольше всего длится инкубационный период при укусе ноги. Выделяют 3 стадии болезни: I — начальную, II — возбуждения, III — паралитическую. Первая стадия начинается с общего недомогания, головной боли, небольшого повышения температуры тела, мышечных болей, сухости во рту, снижения аппетита, болей в горле, сухого кашля, может быть тошнота и рвота. В месте укуса появляются неприятные ощущения — жжение, покраснение, тянущие боли, зуд, повышенная чувствительность. Больной подавлен, замкнут, отказывается от еды, у него возникает необъяснимый страх, тоска, тревога, депрессия, реже — повышенная раздражительность. Характерны также бессонница, кошмары, обонятельные и зрительные галлюцинации. Через 1-3 дня у больного бешенством наступает вторая стадия — возбуждения. Появляется беспокойство, тревога, и, самое характерное для этой стадии, приступы водобоязни. При попытке питья, а вскоре даже при виде и звуке льющейся воды, появляется чувство ужаса и спазмы мышц глотки и гортани. Дыхание становиться шумным, сопровождается болью и судорогами. На этой стадии заболевания человек становится раздражительным, возбудимым, очень агрессивным, «бешеным». Во время приступов больные кричат и мечутся, могут ломать мебель, проявляя нечеловеческую силу, кидаться на людей. Отмечается повышенное пото— и слюноотделение, больному сложно проглотить слюну и он постоянно ее сплевывает. Этот период обычно длится 2-3 дня. Далее наступает третья стадия заболевания, для начала которой характерно успокоение — исчезает страх, приступы водобоязни, возникает надежда на выздоровление. После этого повышается температура тела свыше 40-42°С, наступает паралич конечностей, нарушения сознания, судороги. Смерть наступает от паралича дыхания или остановки сердца. Таким образом, продолжительность заболевания редко превышает неделю. Иногда заболевание без предвестников сразу начинается со стадии возбуждения или появления параличей. У детей бешенство характеризуется более коротким инкубационным периодом. Приступы гидрофобии и резкого возбуждения могут отсутствовать. Заболевание проявляется депрессией, сонливостью, развитием параличей и коллапса. Смерть может наступить через сутки после начала болезни. В качестве вариантов течения выделяют бульбарные, паралитические (типа Ландри), менингоэнцефалитические и мозжечковые формы болезни.  Прогноз Неблагоприятный. Имеется лишь несколько случаев выздоровления. Лабораторная диагностика 1. Вирусоскопический метод. Обнаружение телец Бабеша-Негри в клетках аммониевого рога (используется для посмертной диагностики). 2. Вирусологический метод. Выделение вируса из слюны больных, взвеси мозговой ткани или подчелюстных слюнных желез умерших путем заражения мышей (интрацеребрально) или хомяков (внутрибрюшинно), а также в культуре тканей. 3. Иммунофлуоресцентный метод. Исследуют срезы мозговой ткани, обработанные специфической люминесцирующей сывороткой, с целью обнаружения АГ вируса бешенства. Реакция иммунофлюоресценции, несмотря на то, что применяется уже более 50 лет, остается золотым стандартом в диагностике бешенства, так как обладает рядом достоинств: высокая чувствительность и специфичность; быстрота выполнения (2-3 часа) и относительная дешевизна. Но существуют и недостатки: отсутствие инструментального учета; субъективность оценки результатов, в связи с чем необходимо профессиональное обучение и приобретение опыта работы. Лечение В конце XIX века (1881 г.), работая в области иммунологии, Луи Пастер получил вакцину бешенства. В 1885 г. Л. Пастер впервые применил вакцину на мальчике, укушенном собакой, известен случай и российских крестьян, которые явились к нему в лабораторию из царской России во Францию. Эффективных методов лечения пока нет. Применяют симптоматическое лечение, имеющего целью защитить больного от внешних раздражителей, поддержать водный баланс. Двигательное возбуждение снимают успокаивающими (седативными) средствами, судороги устраняют курареподобными препаратами. Дыхательные расстройства компенсируют посредством трахеотомии и подключения больного к аппарату искусственного дыхания. Больного помещают в отдельную палату, максимально ограждают от различных раздражителей(шум, свет, пыль, движения воздуха, яркий свет). За больными устанавливается постоянное наблюдение. В 2005 году появились сообщения о том, что 15-летняя девушка из США Джина Гис смогла выздороветь после заражения вирусом бешенства без вакцинации, когда лечение было начато уже после появления клинических симптомов. http://ru.wikipedia.org/wiki/%D0%91%D0%B5%D1%88%D0%B5%D0%BD%D1%81%D1%82%D0%B2%D0%BE — cite_note-7 При лечении Гис была введена в искусственную кому, и затем ей были введены препараты, стимулирующие иммунную активность организма. Метод базировался на предположении, что вирус бешенства не вызывает необратимых поражений центральной нервной системы, а вызывает лишь временное расстройство её функций, и, таким образом, если временно «отключить» большую часть функций мозга, то организм постепенно сможет выработать достаточное количество антител, чтобы победить вирус. После недели нахождения в коме и последующего лечения Гис через несколько месяцев была выписана из больницы без признаков поражения вирусом бешенства. [9] Тем не менее, все последующие попытки использовать тот же метод на других пациентах не привели к успеху. Среди медиков до сих пор не прекращаются дискуссии о том, почему выздоровела Джина Гис. Некоторые указывают на то, что она могла быть заражена сильно ослабленной формой вируса или имела необычайно сильный иммунный ответ. Третьим в мире подтверждённым случаем, когда человеку удалось выздороветь от бешенства без использования вакцины, является факт излечения 15-летнего подростка, госпитализированного с симптомами бешенства в Бразилии. http://ru.wikipedia.org/wiki/%D0%91%D0%B5%D1%88%D0%B5%D0%BD%D1%81%D1%82%D0%B2%D0%BE — cite_note-medportal.ru-3 Подросток, имя которого не раскрывается, заразился бешенством в результате укуса летучей мыши в бразильском штате Пернамбуко. По неизвестным причинам мальчику не проводилась вакцинация, позволяющая избежать развития заболевания. В октябре у ребёнка появились симптомы поражения нервной системы, характерные для бешенства, и он был госпитализирован в Университетскую больницу Освальдо Круза (Osvaldo Cruz University Hospital) в Ресифе, столице штата Пернамбуко (Бразилия). Для лечения мальчика врачи использовали комбинацию противовирусных препаратов, седативных средств и инъекционных анестетиков. По словам лечащих врачей, спустя месяц после начала лечения в крови мальчика вирус отсутствовал. Ребёнок пошёл на поправку. Профилактика Госпитализация. Обязательна. Изоляция контактных. Не производится. За укусившими животными устанавливается наблюдение в течение 10 дней. Бешеных и подозрительных на бешенство животных уничтожают и направляют их головной мозг для лабораторного исследования.
Прогноз Неблагоприятный. Имеется лишь несколько случаев выздоровления. Лабораторная диагностика 1. Вирусоскопический метод. Обнаружение телец Бабеша-Негри в клетках аммониевого рога (используется для посмертной диагностики). 2. Вирусологический метод. Выделение вируса из слюны больных, взвеси мозговой ткани или подчелюстных слюнных желез умерших путем заражения мышей (интрацеребрально) или хомяков (внутрибрюшинно), а также в культуре тканей. 3. Иммунофлуоресцентный метод. Исследуют срезы мозговой ткани, обработанные специфической люминесцирующей сывороткой, с целью обнаружения АГ вируса бешенства. Реакция иммунофлюоресценции, несмотря на то, что применяется уже более 50 лет, остается золотым стандартом в диагностике бешенства, так как обладает рядом достоинств: высокая чувствительность и специфичность; быстрота выполнения (2-3 часа) и относительная дешевизна. Но существуют и недостатки: отсутствие инструментального учета; субъективность оценки результатов, в связи с чем необходимо профессиональное обучение и приобретение опыта работы. Лечение В конце XIX века (1881 г.), работая в области иммунологии, Луи Пастер получил вакцину бешенства. В 1885 г. Л. Пастер впервые применил вакцину на мальчике, укушенном собакой, известен случай и российских крестьян, которые явились к нему в лабораторию из царской России во Францию. Эффективных методов лечения пока нет. Применяют симптоматическое лечение, имеющего целью защитить больного от внешних раздражителей, поддержать водный баланс. Двигательное возбуждение снимают успокаивающими (седативными) средствами, судороги устраняют курареподобными препаратами. Дыхательные расстройства компенсируют посредством трахеотомии и подключения больного к аппарату искусственного дыхания. Больного помещают в отдельную палату, максимально ограждают от различных раздражителей(шум, свет, пыль, движения воздуха, яркий свет). За больными устанавливается постоянное наблюдение. В 2005 году появились сообщения о том, что 15-летняя девушка из США Джина Гис смогла выздороветь после заражения вирусом бешенства без вакцинации, когда лечение было начато уже после появления клинических симптомов. http://ru.wikipedia.org/wiki/%D0%91%D0%B5%D1%88%D0%B5%D0%BD%D1%81%D1%82%D0%B2%D0%BE — cite_note-7 При лечении Гис была введена в искусственную кому, и затем ей были введены препараты, стимулирующие иммунную активность организма. Метод базировался на предположении, что вирус бешенства не вызывает необратимых поражений центральной нервной системы, а вызывает лишь временное расстройство её функций, и, таким образом, если временно «отключить» большую часть функций мозга, то организм постепенно сможет выработать достаточное количество антител, чтобы победить вирус. После недели нахождения в коме и последующего лечения Гис через несколько месяцев была выписана из больницы без признаков поражения вирусом бешенства. [9] Тем не менее, все последующие попытки использовать тот же метод на других пациентах не привели к успеху. Среди медиков до сих пор не прекращаются дискуссии о том, почему выздоровела Джина Гис. Некоторые указывают на то, что она могла быть заражена сильно ослабленной формой вируса или имела необычайно сильный иммунный ответ. Третьим в мире подтверждённым случаем, когда человеку удалось выздороветь от бешенства без использования вакцины, является факт излечения 15-летнего подростка, госпитализированного с симптомами бешенства в Бразилии. http://ru.wikipedia.org/wiki/%D0%91%D0%B5%D1%88%D0%B5%D0%BD%D1%81%D1%82%D0%B2%D0%BE — cite_note-medportal.ru-3 Подросток, имя которого не раскрывается, заразился бешенством в результате укуса летучей мыши в бразильском штате Пернамбуко. По неизвестным причинам мальчику не проводилась вакцинация, позволяющая избежать развития заболевания. В октябре у ребёнка появились симптомы поражения нервной системы, характерные для бешенства, и он был госпитализирован в Университетскую больницу Освальдо Круза (Osvaldo Cruz University Hospital) в Ресифе, столице штата Пернамбуко (Бразилия). Для лечения мальчика врачи использовали комбинацию противовирусных препаратов, седативных средств и инъекционных анестетиков. По словам лечащих врачей, спустя месяц после начала лечения в крови мальчика вирус отсутствовал. Ребёнок пошёл на поправку. Профилактика Госпитализация. Обязательна. Изоляция контактных. Не производится. За укусившими животными устанавливается наблюдение в течение 10 дней. Бешеных и подозрительных на бешенство животных уничтожают и направляют их головной мозг для лабораторного исследования.
Предупреждение бродяжничества собак и кошек, профилактическая иммунизация домашних животных, тщательная первичная обработка укушенных ран. Для успешной борьбы с бешенством диких животных необходимо «изъятие» восприимчивых животных из популяции. Это можно осуществить путем контроля популяции животных и путем оральной иммунизации Людям, укушенным бешеными или неизвестными животными, местную обработку раны необходимо проводить немедленно или как можно раньше после укуса или повреждения; рану обильно промывают водой с мылом (или детергентом) и обрабатывают 40-70 градусным спиртом или настойкой йода, при наличии показаний вводят антирабический иммуноглобулин вглубь раны и в мягкие ткани вокруг неё.
1. Сухие антирабические вакцины типа Ферми и КАВ применяют для активной иммунизации по условным и безусловным показаниям. Показания к проведению прививок, доза вакцины и длительность курса иммунизации определяются врачами, получившими специальную подготовку. 2. Антирабический иммуноглобулин из сыворотки лошади применяют с целью создания немедленного пассивного иммунитета. Вакцины, используемые в настоящее время, как правило, вводятся 6 раз: инъекции делаются в день обращения к врачу (0 день), а затем на 3, 7, 14, 30 и 90 дни. Если за укусившим животным удалось установить наблюдение, и в течение 10 суток после укуса оно осталось здоровым, то дальнейшие инъекции прекращают. Во время вакцинации и в течение 6 месяцев после последней прививки запрещено употребление алкоголя. В период вакцинации также необходимо ограничить употребление в пищу продуктов, способных вызвать у пациента аллергическую реакцию.
Для продолжения скачивания необходимо пройти капчу:
Сервис бесплатной оценки стоимости работы
- Заполните заявку. Специалисты рассчитают стоимость вашей работы
- Расчет стоимости придет на почту и по СМС
Прямо сейчас на почту придет автоматическое письмо-подтверждение с информацией о заявке.
источник
 Бешенство Бешенство практически во всех случаях является фатальным заболеванием, которое может манифестировать как классическая буйная или как паралитическая форма. Выздоровление описано всего у нескольких больных, большинство из которых были инфицированы вариантом вируса бешенства летучих мышей, и сопровождалось быстрым иммунным ответом инфицированного организма и самопроизвольной (иммунной) элиминацией вируса. В целом удовлетворительное функциональное восстановление выживших может быть обусловлено эволюцией вирусных механизмов в сторону минимизации повреждения ЦНС, но при этом позволяющих вирусу распространяться. Худшая выживаемость больных с буйной формой по сравнению с показателем у пациентов с паралитической тесно соотносится с большим количеством вируса и более слабым иммунным ответом в ЦНС при буйной форме. Вирус бешенства появляется в ЦНС задолго до возникновения симптомов — у больных с буйной формой еще за несколько дней до развития церебральной симптоматики по данным МРТ головного мозга можно выявить субклиническую дисфункцию клеток переднего рога и признаки поражения. Мы только сейчас начинаем понимать, посредством каких механизмов реализуется разрушительное действие вируса бешенства, каким образом он селективно поражает функцию периферических нервов у пациентов с паралитической формой и нарушает поведение у больных с буйной формой бешенства. Однако для разработки эффективного лечения необходимо детальное понимание нейропатогенетических механизмов.
Бешенство Бешенство практически во всех случаях является фатальным заболеванием, которое может манифестировать как классическая буйная или как паралитическая форма. Выздоровление описано всего у нескольких больных, большинство из которых были инфицированы вариантом вируса бешенства летучих мышей, и сопровождалось быстрым иммунным ответом инфицированного организма и самопроизвольной (иммунной) элиминацией вируса. В целом удовлетворительное функциональное восстановление выживших может быть обусловлено эволюцией вирусных механизмов в сторону минимизации повреждения ЦНС, но при этом позволяющих вирусу распространяться. Худшая выживаемость больных с буйной формой по сравнению с показателем у пациентов с паралитической тесно соотносится с большим количеством вируса и более слабым иммунным ответом в ЦНС при буйной форме. Вирус бешенства появляется в ЦНС задолго до возникновения симптомов — у больных с буйной формой еще за несколько дней до развития церебральной симптоматики по данным МРТ головного мозга можно выявить субклиническую дисфункцию клеток переднего рога и признаки поражения. Мы только сейчас начинаем понимать, посредством каких механизмов реализуется разрушительное действие вируса бешенства, каким образом он селективно поражает функцию периферических нервов у пациентов с паралитической формой и нарушает поведение у больных с буйной формой бешенства. Однако для разработки эффективного лечения необходимо детальное понимание нейропатогенетических механизмов.
Бешенство, заболевание которым вызывает вирус бешенства (rabies virus — RABV) 1-го генотипа, является одной из наиболее распространенных фатальных инфекций во всем мире. В Европе, Азии и Африке это заболевание в основном ассоциируется с укусами собак, в Америке — с укусами летучих мышей. У 2/3 инфицированных собачьими вариантами RABV манифестируют классической буйной формой бешенства, которая характеризуется флуктуацией уровня сознания, изменением психического статуса, фобическими или инспираторными спазмами и симптомами раздражения вегетативной нервной системы. У 1/3 больных развивается паралитическая форма бешенства, которая напоминает синдром Гийена — Барре, однако нарастание комы, отек мышц и недержание мочи позволяют с уверенностью различать эти заболевания (таблица). Бешенство, ассоциированное с вариантами RABV летучих мышей, имеет такие атипичные черты, как фокальные симптомы поражения ствола мозга, миоклонус, гемихорея, симптомы и признаки синдрома Горнера.
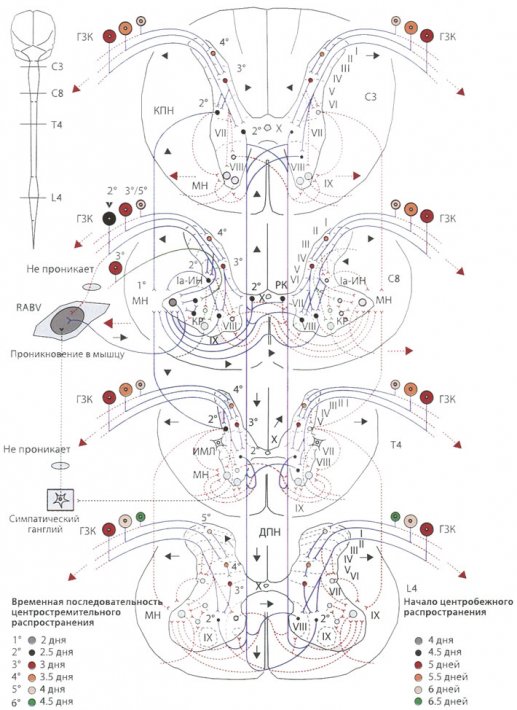 Пути распространения RABV к и по спинному мозгу
Пути распространения RABV к и по спинному мозгу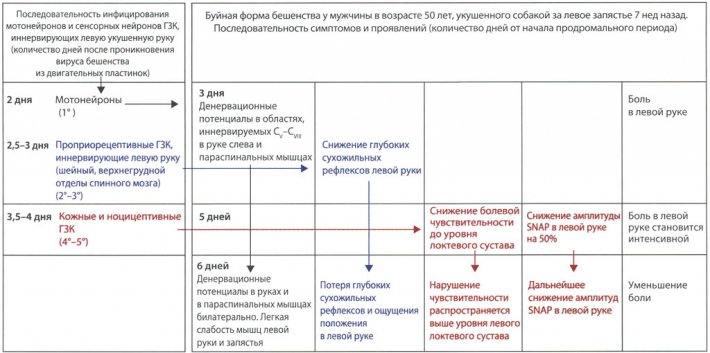 Связь между продромальными симптомами и проявлениями и распространением вируса бешенства по спинному мозгу у больного с буйной формой бешенства
Связь между продромальными симптомами и проявлениями и распространением вируса бешенства по спинному мозгу у больного с буйной формой бешенства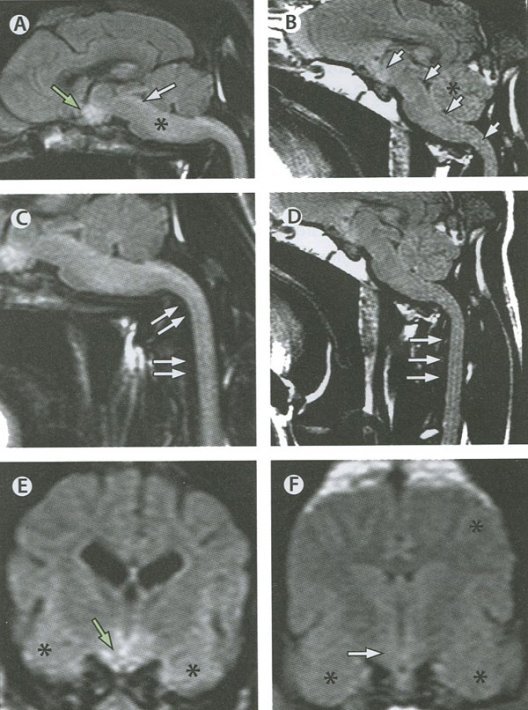 МРТ при собачьем бешенстве
МРТ при собачьем бешенстве